Основные свойства живого
Вспомним, что сущность теории Дарвина состоит в появлении небольших изменений в генетическом материале и, в результате, в организме. В то время механизм этих изменений еще не был известен. Мы уже говорили, что избыточность генетического кода допускает некоторые изменения нуклеотидной последовательности без изменений в кодируемых белковых продуктах.

Кроме того, аминокислоты, кодируемые подобными последовательностями, структурно похожи друг на друга. Далее, формирование конечных трехмерных белковых продуктов допускает такие изменения в коде, что созданные белки получаются не идентичными, но похожими и могут выполнять те же функции. Если аминокислота в белке заменена на другую, имеющую похожие химические свойства (тот же заряд или полярность), то в полипептидной цепочке она будет вести себя так же, как исходная, и создавать те же или похожие объемные структуры, которые будут выполнять те же функции. Поскольку именно функция белков является наиболее важным их свойством, которое необходимо сохранить, последовательности могут в некоторой степени изменяться при передаче генов от поколения к поколению, но трехмерная структура и функции генных продуктов (белков) остаются неизменными. На основании неизменных свойств ген, унаследованный от общего предка двумя линиями потомков, может быть признан как родственный (гомологичный) ген этих двух линий, хотя в каждой из них в нем накопился определенный уровень мутаций с момента отхода от общего предка. Так видоизменяются постулаты Дарвина.
Количество мутаций у двух родственных видов напрямую связано со временем, в течение которого они эволюционировали врозь, так что это различие можно использовать для статистических измерений родственных связей между двумя видами. Эти связи удобно изобразить в виде филогенетических деревьев, где ветви представляют родственные связи организмов, а длина ветвей указывает на генетическое расстояние (расхождение) между видами после их разделения. Большинство мутаций либо незначительны, либо вредны, но некоторые действительно способствуют выживанию. Таково современное представление о корневом механизме эволюции.
Все упомянутые выше основные свойства жизни, такие как структура и состав генетического материала, генетический код и 20 аминокислот, используемых во всех белках, а также аппарат трансляции, почти совершенно идентичны у всех видов, живущих сегодня на Земле. Очевидно, эти свойства были приобретены еще на ранних стадиях эволюции и, повидимому, имелись у последнего общего предка в то время, когда из него выделились три ветви жизни. Связь
Всех этих форм жизни можно исследовать путем сравнения последовательностей тех РНК (например, 168 РНК), которые участвуют в трансляции — одном из наиболее эволюционно консервативных процессов. На основании последовательности 168 РНК первое филогенетическое дерево построили в 1990 году Карл Вёзе и его коллеги. Это дерево показывает, что сначала две линии прокариотов отделились от популяции последнего общего предка, а затем от них отделились эукариоты.
Заметим, что позже филогенетическое дерево и порядок происхождения трех его ветвей были поставлены под вопрос, так как по другим последовательностям генов были получены другие филогенетические связи между многими организмами. Эти различия можно объяснить тем, что многие гены обменивались между разными видами уже после их разделения по разным филогенетическим линиям, а также тем, что эволюцию разных генов, повидимому, невозможно сравнивать на больших отрезках времени, поскольку она происходит с разной скоростью.
Кроме упомянутых выше генетических элементов жизни, еще одним вездесущим свойством жизни, берущим свое начало у очень ранних ее форм, является гомохиралъностъ «строительных блоков» нуклеиновых кислот и белков. Хиральность — это свойство молекул, имеющих атом (например, углерод) с четырьмя разными связями. Так как эти связи направлены в четыре разных угла тетраэдра, структура молекулы несимметрична, и ее можно построить в двух разных ориентациях, являющихся зеркальными отражениями друг друга. В нашу эпоху связи центрального (а) углерода во всех аминокислотах несимметричны так же как и связи углерода номер 4 в рибозе и дезоксирибозе Сахаров
В искусственно синтезированных аминокислотах и сахарах эти молекулы с углеродными связями, направленными и в ту и в другую сторону, обычно встречаются в равном количестве (поэтому они не гомохиральны). Но все биогенные аминокислоты гомохиральны и имеют ауглеродные связи в Ьконфигурации (Ь = 1еуо, левый), и все сахара тоже гомохиральны и имеют связи у 4го углерода в Бконфигурации (Б = сехтго, правый). Происхождение этой особой гомохиральности до сих пор не получило объяснения. Ясно, что гомохиральность по сути является химическим «диктатором», позволяющим связать мономерные звенья в изящный линейный полимер, тогда как обратная хиральность перегибает структуру, закручивая ее в противоположную сторону. Но непонятно, почему и как жизнь выбрала левые аминокислоты и правые сахара. Пока мы лишь начинаем понимать, почему именно 20 аминокислот используются в белках и почему именно эти четыре разных нуклеотида применяются в ДНК и РНК. Сейчас нам кажется, что это мог быть случайный выбор молекул, которые существовали в древней окружающей среде, и этот «случай» закрепился и стал «правилом». Как бы то ни было, сейчас мы знаем несколько важнейших свойств жизни на Земле. Если мы когданибудь найдем жизнь гдето еще, то будет очень интересно узнать, обладает ли она теми же свойствами.
Это покажет, имеет ли обнаруженная жизнь то же происхождение, что и наша, или она возникла самостоятельно.
-
Похожие статьи из категории: Химия жизни
-
 Генетика и эволюция жизни
Генетика и эволюция жизни
Чарлз Дарвин (1809-1882) был первым, кто ясно понял, что новые виды формируются в результате генетических изменений и что движущая сила эволюции — это естественный отбор. Дарвин только что окончил Кембриджский университет, когда его пригласили участвовать в многолетней экспедиции на корабле «Бигль», отправлявшемся в кругосветное путешествие. Кульминационным моментом экспедиции Дарвина стало посещение Галапагосских островов, где он обнаружил много новых
-
 Генетический код и его экспрессия
Генетический код и его экспрессия
Генетический код хранится в нуклеотидной последовательности ДНК в форме следующих один за другим триплетов нуклеотидову причем каждый из триплетов соответствует определенной аминокислоте. Порядок триплетов, или ген, показывает, в какой последовательности должны объединиться аминокислоты, чтобы сформировать определенный белок. Используя триплеты, составленные из четырех разных
-
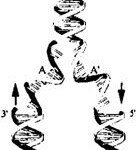 Молекула ДНК
Молекула ДНК
Итак, молекула ДНК имеет две магистральные цепи, обвивающие друг друга и образующие знаменитую «двойную спираль». Основания показаны в виде букв на ступенях этой «лестницы», соединяющих магистрали обеих нитей в единую спираль. Для воспроизводства генетической информации нуклеотидная последовательность ДНК копируется (реплицируется) на параллельную нить, образующую затем с исходной нитью двойную спираль. Интересно, что вторая цепь двойной спирали идет не в том же направлении,
-
 Открытие генетики и ее химические основы
Открытие генетики и ее химические основы
Австрийский монах Грегор Мендель (1822-1884) проделал основополагающую работу для определения законов и механизмов наследственности. В течение многих лет на монастырском огороде он выращивал горох и детально описал все выращенных им растений. При этом он смог проследить в течение нескольких поколений характер наследования
-
 Химия жизни
Химия жизни
Белки — основная рабочая сила и структурный материал клетки. Они имеют различную форму и размер, и каждый из них выполняет в клетке свою задачу. Некоторые белки формируют основные клеточные структуры, такие как нитевидный цитоскелет — каркас клетки, а также компоненты клеточных стенок и разнообразные «ворота» и «насосы» в клеточной мембране. Другой важнейшей функцией различных белков является их работа в качестве химических сигналов и регуляция